

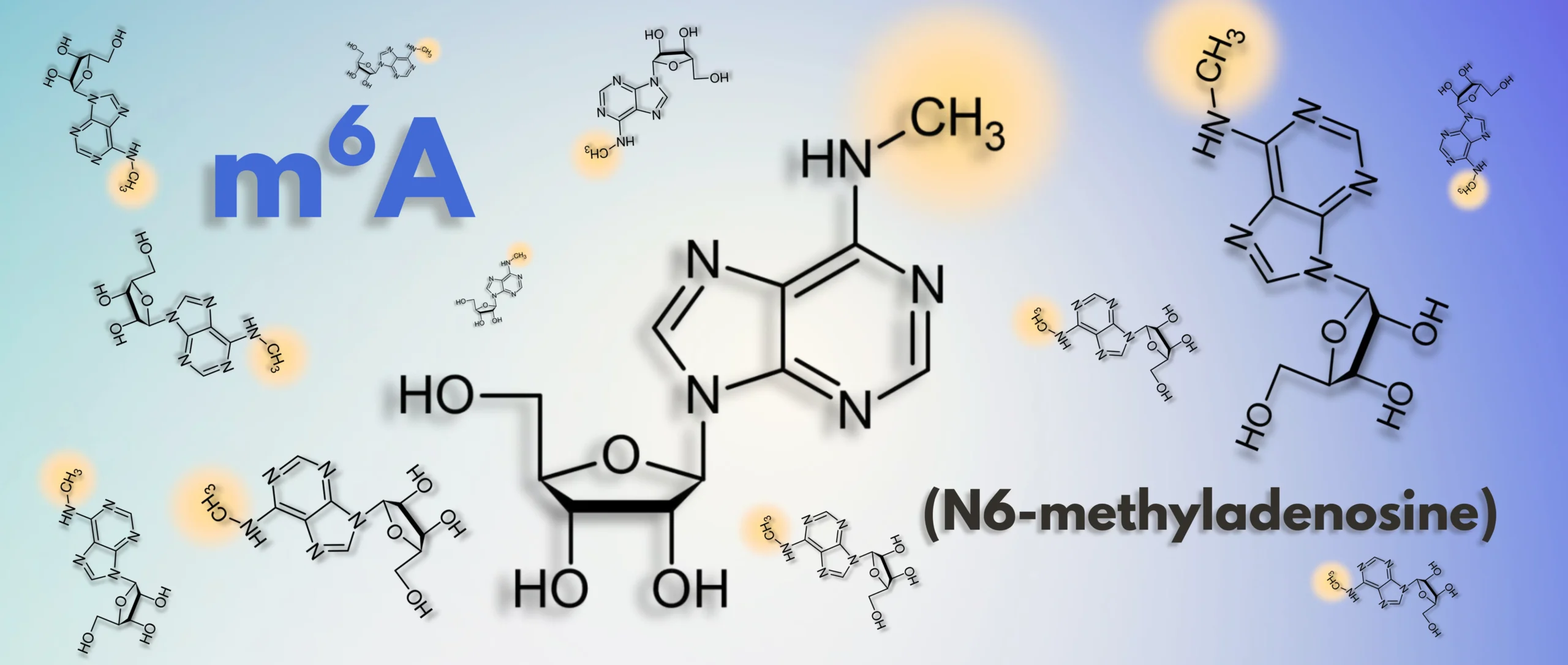

The Hidden Code of RNA
In the world of biology, the central dogma – DNA transcribed into RNA translated into protein – has long been a cornerstone of how we understand life. But in recent years, researchers have discovered a layer of gene regulation that operates beyond this canonical framework: the epitranscriptome. And at the heart of this field is a chemical modification that’s transforming our understanding of RNA regulation and cellular function – N6-methyladenosine, or m6A.
A Quick Intro to m6A
As detailed in a previous post, m6A is the most prevalent internal modification in eukaryotic mRNA occurring at approximately 3 to 5 sites per transcript and accounting for 0.1% to 0.4% of all adenosines.1 Rather than altering the RNA sequence, m6A functions as a dynamic epitranscriptomic signal – akin to a molecular ‘annotation’ – that guides the post-transcriptional fate of RNA molecules. These modifications regulate key processes such as alternative splicing, nuclear export, translation efficiency, and mRNA decay. Enrichment of m6A near stop codons and in 3′ untranslated regions (3′ UTRs) supports its widespread role in regulating mRNA stability and translation.1 This regulatory capacity enables m6A to fine-tune gene expression in a temporal and context-dependent manner, positioning it as a central player in developmental programming, stress responses, and disease states.
To execute its regulatory functions, m6A relies on a biological toolkit composed of ‘writers,’ ‘readers,’ and ‘erasers.’ The writers, such as the methyltransferase complex containing METTL3/METTL14, catalyze the installation of the methyl group onto specific adenosine residues within RNA transcripts. Readers, mainly comprised of the YTH domain-containing families, serve as interpreters of this modification, directing marked RNAs toward distinct fates such as enhanced translation, targeted decay, or alternative splicing. In parallel, erasers, including FTO and ALKBH5, demethylate m6A back to adenosine enabling a reversible and finely tunable mode of regulation. This dynamic interplay between deposition, recognition, and removal of m6A allows for responsive modulation of gene expression in processes ranging from stem cell differentiation to neuronal adaptation.
A three-part toolkit enforces m6A-mediated regulation:
| Role | Key Players | Job Description |
|---|---|---|
| Writers | METTL3-METTL14 complex | Install the methyl groups |
| Readers | YTH-domain proteins | Interpret the mark and route the RNA |
| Erasers | FTO, ALKBH5 | Remove the mark for reversibility |
m6A and Alternative Splicing: Editing the Editor
Among its many regulatory functions, m6A plays a critical role in modulating RNA splicing—a process that expands transcript diversity and fine-tunes gene expression. m6A can influence the recruitment of splicing factors to pre-mRNA and shape exon inclusion or skipping. These effects are primarily mediated by nuclear m6A readers like YTHDC1, which can promote exon inclusion by recruiting the splicing factor SRSF3 while inhibiting binding of SRSF10, a splicing repressor.2 Mechanistically, the presence of m6A near splice junctions can alter RNA secondary structure, exposing or occluding splicing enhancer motifs. This structural change, sometimes referred to as the “m6A switch,” can modulate the accessibility of other RNA-binding proteins like HNRNPC, further influencing splicing outcomes.
In human disease, the regulation of m6A has been identified as a hallmark of Huntington’s disease (HD),3 and a recent study detailed a specific role for m6A in alternative splicing and HD progression. The study by Nguyen et al. revealed that in HD, there is a reduction in m6A RNA modification on transcripts abnormally expressed in the striatum—a critical region of the brain involved in motor control and cognitive function.4 This hypomethylation occurs at sites adjacent to TDP-43 binding regions. TDP-43, a critical RNA-binding protein, exhibits mislocalization and loss of function in HD, leading to aberrant splicing events. The study suggests that the combined effect of diminished m6A modification and TDP-43 dysfunction contributes to the splicing abnormalities observed in HD.
A complementary new report from Pupak et al. (2024) further illuminates how m6A contributes to HD pathology by showing increased methylation within intron 1 of mutant HTT transcripts at cryptic polyadenylation sites, which enhances the production of toxic HTT1a splice variants in both mouse models and human striatal tissue.5 These examples underscore the intricate interplay between m6A modifications and RNA-binding proteins in maintaining proper splicing regulation, and how their disruption can lead to disease pathogenesis.
A compelling example of m6A’s influence on splicing in cancer biology comes from a recent study by Achour et al., which investigates the role of METTL3 in breast tumorigenesis. The authors demonstrate that METTL3 is upregulated in breast cancer cell lines and that its silencing impairs cell proliferation and promotes apoptosis.6 Strikingly, METTL3 depletion also induces widespread alternative splicing (AS) changes, particularly exon skipping and alternative first exon events, many of which are enriched in genes associated with cell adhesion, apoptosis, and splicing itself. Ultimately, the study shows that METTL3 promotes splicing of cancer-associated transcripts through both direct and indirect routes: m6A modifications are enriched near splice junctions of target genes, and METTL3 regulates the expression of splicing regulators, notably through m6A-mediated control of MYC. Remarkably, METTL3-regulated AS events were correlated with poor prognosis in breast cancer patients and showed subtype-specific expression patterns, suggesting their utility as potential biomarkers. This work highlights how m6A serves as a nexus between transcriptional regulation and splicing control in cancer, adding a new layer to our understanding of how tumors co-opt the epitranscriptome to support malignant progression.
One biologically significant example of m6A-mediated splicing regulation comes from studies of sex determination in Drosophila. It powerfully illustrates the evolutionary conservation and developmental relevance of m6A’s role in alternative splicing. In flies, m6A marks on the Sex-lethal (Sxl) pre-mRNA regulate the inclusion of a key exon required for female-specific isoforms.7 The reader protein YT521-B (the Drosophila ortholog of reader protein YTHDC1) binds m6A sites and ensures the proper splicing pattern for female development. Loss of m6A deposition leads to aberrant Sxl splicing and a switch to male-specific isoforms, resulting in phenotypic sex reversal.
The “m6A switch” recruits splicing factors, and where needed, reshape RNA structure to expose or hide splicing motifs
| Neurodegeneration | In Huntington’s disease, hypomethylation appears beside TDP-43 binding sites, compounding TDP-43 loss-of-function and driving aberrant splicing |
| Cancer | In breast tumors, METTL3 over-expression fuels proliferation and rewires hundreds of splice events—many tied to poor prognosis—by decorating key junctions and controlling splicing-factor transcripts like MYC |
| Development | In Drosophila, m6A marks on Sex-lethal pre-mRNA ensure female-specific exon inclusion; erasing them flips flies toward male isoforms |
Timing Is Everything: m6A and RNA Life Cycles
Much of what makes m6A biologically significant boils down to timing. By methylating RNA transcripts, cells can regulate both the rate of mRNA degradation and translation. This precision gives cells the ability to rapidly shift their gene expression programs—critical during developmental transitions, responses to environmental stress, or rapid cell signaling.
During the maternal-to-zygotic transition in early embryos, m6A helps clear out maternal mRNAs so that the zygotic genome can take control.8 In immune cells, m6A plays a multifaceted role in shaping immune responses. It controls the decay of pro-inflammatory cytokine mRNAs, thereby tuning the magnitude and duration of inflammatory signaling.9 m6A also influences the development and differentiation of various immune cell types. For instance, deletion of METTL3 in T cells impairs their proliferation into effector or regulatory subtypes, with downstream consequences on immune homeostasis and response. Similarly, in macrophages, m6A regulates polarization states by modulating transcript stability for inflammatory or anti-inflammatory programs. These findings position m6A as a key regulator of immune cell plasticity, ensuring proper immune surveillance, resolution of inflammation, and prevention of autoimmunity.
In essence, m6A provides a temporal switch. Want to shut off a pluripotency program? Add m6A to those mRNAs. Need to boost protein synthesis during learning? Use m6A to enhance translation. These decisions play out in milliseconds within cells and have far-reaching effects on tissue development and function.
m6A Functions
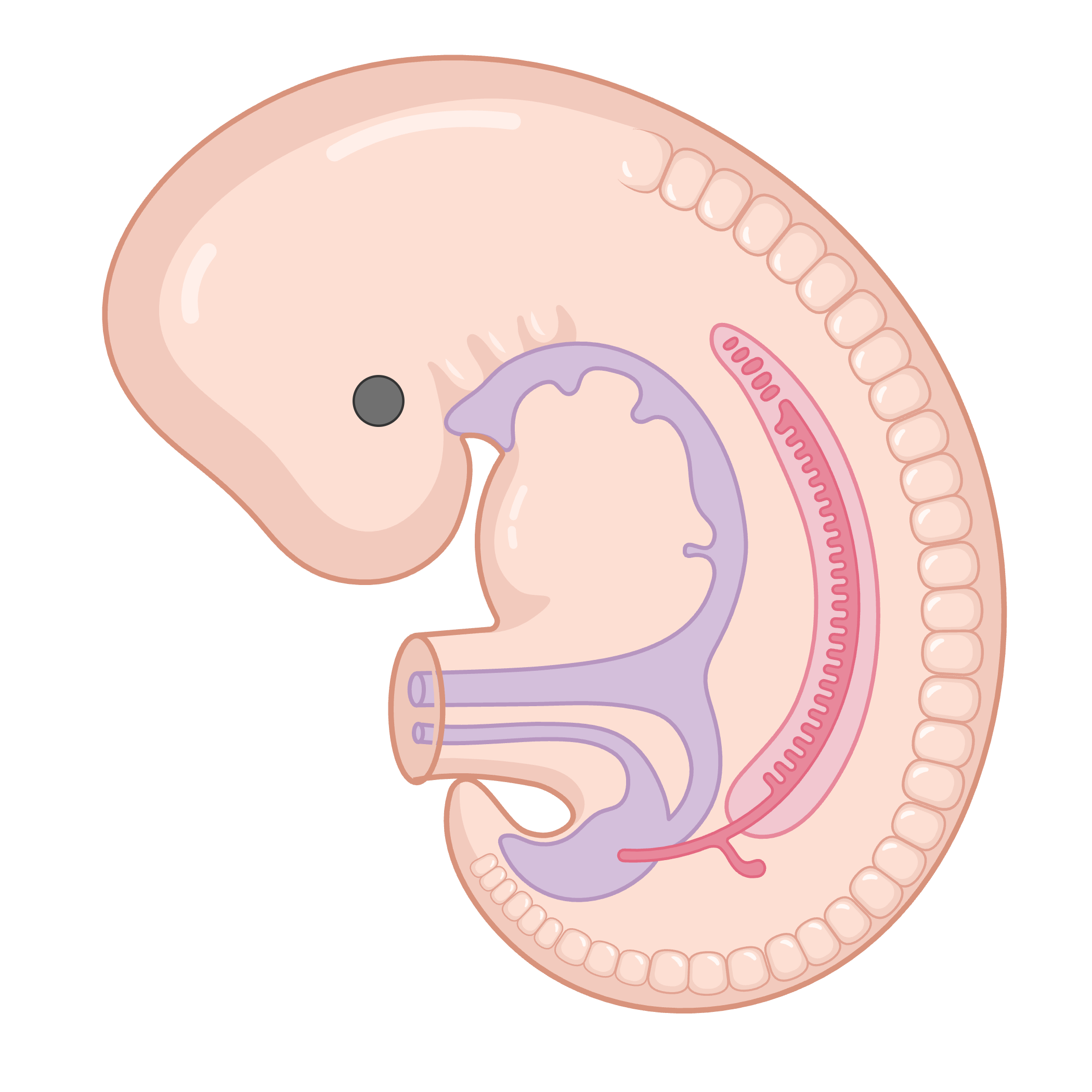
m6A marks maternal mRNAs for degradation so zygotic genes can assume control.
m6A tempers the intensity and duration of inflammation by shortening the half-life of pro-inflammatory cytokine mRNAs. It also guides lineage decisions in T cells and macrophages.
Adding m6A to pluripotency genes like Nanog and Sox2 marks their transcripts for degradation, letting stem cells exit self‑renewal.
Writing the Script for Development
One of the clearest examples of m6A’s importance is during embryonic development. Without it, stem cells fail to differentiate. In mouse embryonic stem cells, depletion of the methyltransferase enzymes METTL3 and METTL14 that install m6A results in a frozen state where cells continue to self-renew but cannot progress toward specialized identities.10,11 Methylation by m6A helps turn off key pluripotency factors (like Nanog and Sox2) by marking their mRNAs for degradation, thus allowing differentiation to proceed.
This regulation doesn’t stop at the embryo. m6A also fine-tunes adult stem cell behavior. In hematopoietic stem cells, multipotent progenitors residing in the bone marrow that give rise to all blood and immune cell types, m6A helps balance self-renewal with the need to differentiate into various blood lineages. Without proper methylation, blood stem cells either overproliferate or fail to produce the right mix of mature immune cells. m6A accomplishes this by controlling the stability of mRNAs tied to key signaling pathways like Notch and Wnt.12-14
Rewiring the Brain, One Transcript at a Time
Nowhere is m6A’s influence more striking than in the brain. The central nervous system has some of the highest levels of m6A methylation in the body, and for good reason. From neurogenesis to synaptic plasticity, m6A guides the brain’s development and ongoing function.
In developing neurons, m6A directs the timely exit of neural stem cells from the cell cycle so they can begin differentiating into functional neurons and glial cells.15 Later, during processes like myelination, m6A ensures that oligodendrocyte precursor cells mature properly to insulate axons and support neuronal communication.16
But the role of m6A doesn’t end at development—it’s also active in learning and memory. In the adult brain, m6A boosts the translation of synaptic proteins in response to stimuli, supporting long-term memory formation.17 Mouse models lacking METTL3 or YTHDF1 show clear deficits in memory consolidation, underscoring how vital this regulation is to higher-order brain functions.18
m6A at the Crossroads of Disease
Given its central role in gene regulation, it’s not surprising that m6A has been implicated in a wide range of human diseases. Its machinery is frequently dysregulated in cancer, where altered m6A patterns support tumor cell proliferation, survival, and immune evasion. In some cases, the enzymes that remove m6A marks (like FTO) are upregulated, helping cancer cells stabilize oncogenic transcripts.19 In others, overactive writers like METTL3 boost the translation of growth-promoting genes.20
Beyond cancer, m6A has also been linked to neurological and psychiatric conditions. In mouse models, deficits in m6A regulators impair neurogenesis and lead to cognitive dysfunction.1 There’s growing evidence that m6A patterns are altered in Alzheimer’s disease, depression, and stress-related disorders, where they may influence how the brain responds to injury or prolonged emotional stress.21
m6A also plays a role in infectious disease. Many viruses—like HIV and SARS-CoV-2—interact with the host’s m6A machinery to enhance their replication.22,23 Some viruses even co-opt m6A to improve the stability and translation of their own RNAs, effectively hijacking the host’s regulatory systems for their benefit. For example, studies of HIV-1 have shown that viral transcripts are dynamically modified by m6A, which enhances their nuclear export and translation efficiency, thus increasing viral replication efficiency.24
m6A dysregulation surfaces in a broad variety of disorders
| Cancer | Tumors may overexpress erasers like FTO to stabilize oncogenic RNAs or up-regulate writers like METTL3 for growth signals |
| Neurological & psychiatric diseases | Altered m6A landscapes have been reported in Alzheimer’s, depression, and stress disorders |
| Infection | Viruses from HIV to SARS-CoV-2 hijack host m6A machinery to speed their own replication |
A New Frontier in Biomedicine
The discovery of m6A’s role in shaping RNA fate has opened a new frontier in molecular biology. As our ability to map and manipulate these marks improves, researchers are exploring how to harness m6A for therapeutic gain. Inhibitors of FTO, for example, have shown promise in treating certain leukemias by restoring methylation on tumor-suppressive transcripts. Likewise, m6A patterns are emerging as potential biomarkers for disease diagnosis and prognosis.
As mapping and editing tools improve, m6A is moving from curiosity to the clinic
| Therapeutics | METTL3 inhibitors attenuate methylation of tumor-promoting transcripts in certain leukemias |
| Biomarkers | Disease-specific m6A signatures may aid diagnosis and prognosis |
By controlling when, where, and how mRNAs are used, m6A represents a key regulatory hub in human biology. From early development to adult tissue maintenance, and from health to disease, this tiny mark exerts a big impact. Understanding it doesn’t just change how we think about RNA—it changes how we think about life itself.
About the Author
Zachary Miles
Zachary Miles is the Sr. Director of Protein Engineering at AlidaBio. He earned a PhD in biochemistry at the University of Arizona studying biosynthetic pathways and enzymatic mechanisms of RNA modification. A postdoctoral position at The University of California – San Diego exposed him to the extensive biotechnology ecosystem where he then utilized this expertise in protein structure/function to help design enzymes and proteins at multiple companies in the San Diego area.
References
1. Meyer, K.D., Saletore, Y., Zumbo, P., Elemento, O., Mason, C.E., & Jaffrey, S.R. (2012). Comprehensive analysis of mRNA methylation reveals enrichment in 3′ UTRs and near stop codons. Cell, 149(7), 1635–1646. https://doi.org/10.1016/j.cell.2012.05.003
2. Xiao, W., Adhikari, S., Dahal, U., Chen, Y.S., Hao, Y.J., Sun, B.F., … & Yang, Y.G. (2016). Nuclear m6A reader YTHDC1 regulates mRNA splicing. Molecular Cell, 61(4), 507–519. https://doi.org/10.1016/j.molcel.2016.01.012
3. Pupak, A., Singh, A., Sancho-Balsells, A., Alcalá-Vida, R., Espina, M., Giralt, A., … & Brito, V. (2022). Altered m6A RNA methylation contributes to hippocampal memory deficits in Huntington’s disease mice. Cellular and Molecular Life Sciences, 79(8), 416. https://doi.org/10.1007/s00018-022-04444-6
4. Nguyen, T. B.; Miramontes, R.; Chillon‑Marinas, C.; Maimon, R.; Vazquez‑Sanchez, S.; Lau, A. L.; McClure, N. R.; Wu, Z.; Wang, K. Q.; England, W. E.; Singha, M.; Stocksdale, J. T.; Heath, M.; Jang, K.‑H.; Jung, S.; Ling, K.; Jafar‑nejad, P.; McKnight, J. I.; Ho, L. N.; Al Dalahmah, O.; Faull, R. L. M.; Steffan, J. S.; Reidling, J. C.; Jang, C.; Lee, G.; Cleveland, D. W.; Lagier‑Tourenne, C.; Spitale, R. C.; Thompson, L. M. Aberrant splicing in Huntington’s disease accompanies disrupted TDP‑43 activity and altered m⁶A RNA modification. Nat. Neurosci. 2025, 28, 280–292. https://doi.org/10.1038/s41593-024-01850-w.
5. Pupak, A., Rodríguez-Navarro, I., Sathasivam, K., Singh, A., Essmann, A., Del Toro, D., Ginés, S., Pinto, R. M., Bates, G. P., Ørom, U. A. V., Martí, E., & Brito, V. (2024). m6A modification of mutant huntingtin RNA promotes the biogenesis of pathogenic huntingtin transcripts. EMBO Reports, 25(11), 5026–5052. https://doi.org/10.1038/s44319-024-00283-7
6. Achour, C.; Bhattarai, D. P.; Groza, P.; Román, Á.‑C.; Aguiló, F.; et al. METTL3 regulates breast cancer‑associated alternative splicing switches. Oncogene 2023, 42, 911–925. https://doi.org/10.1038/s41388-023-02602-z.
7. Kan, L., Grozhik, A.V., Vedanayagam, J., Patil, D.P., Pang, N., Lim, K.S., … & Lai, E.C. (2017). The m6A pathway facilitates sex determination in Drosophila. Nature Communications, 8, 15737. https://doi.org/10.1038/ncomms15737
8. Zhao, B.S., Wang, X., Beadell, A.V., Lu, Z., Shi, H., Kuuspalu, A., … & He, C. (2017). m6A-dependent maternal mRNA clearance facilitates zebrafish maternal-to-zygotic transition. Nature, 542(7642), 475–478. https://doi.org/10.1038/nature21355
9. Wang, X., Lu, Z., Gomez, A., Hon, G.C., Yue, Y., Han, D., … & He, C. (2014). N6-methyladenosine-dependent regulation of messenger RNA stability. Nature, 505(7481), 117–120. https://doi.org/10.1038/nature12730
10. Batista, P.J., Molinie, B., Wang, J., Qu, K., Zhang, J., Li, L., … & Chang, H.Y. (2014). m6A RNA modification controls cell fate transition in mammalian embryonic stem cells. Cell Stem Cell, 15(6), 707–719. https://doi.org/10.1016/j.stem.2014.09.019
11. Geula, S., Moshitch-Moshkovitz, S., Dominissini, D., Mansour, A.A., Kol, N., Salmon-Divon, M., … & Hanna, J.H. (2015). m6A mRNA methylation facilitates resolution of naïve pluripotency toward differentiation. Science, 347(6225), 1002–1006. https://doi.org/10.1126/science.1261417
12. Lv, J., Zhang, Y., Gao, S., Zhang, C., Chen, Y., Li, W., Yang, Y.G., Zhou, Q., & Liu, F. (2018). Endothelial-specific m6A modulates mouse hematopoietic stem and progenitor cell development via Notch signaling. Cell Research, 28(2), 249–252. https://doi.org/10.1038/cr.2017.143
13. Luo, H.; Cortés-López, M.; Tam, C. L.; Xiao, M.; Wakiro, I.; Chu, K. L.; Pierson, A.; Chan, M.; Chang, K.; Yang, X.; Fecko, D.; Han, G.; Ahn, E. E.; Morris, Q. D.; Landau, D. A.; Kharas, M. G. SON is an essential m⁶A target for hematopoietic stem cell fate. Cell Stem Cell 2023, 30(12), 1658–1673.e10. https://doi.org/10.1016/j.stem.2023.11.006.
14. Ji, R., & Zhang, X. (2021). The roles of m6A in regulating stem cell fate. Frontiers in Cell and Developmental Biology, 9, 765635. https://doi.org/10.3389/fcell.2021.765635
15. Yoon, K.J., Ringeling, F.R., Vissers, C., Jacob, F., Pokrass, M., Jimenez-Cyrus, D., … & Song, H. (2017). Temporal control of mammalian cortical neurogenesis by m6A methylation. Cell, 171(4), 877–889.e17. https://doi.org/10.1016/j.cell.2017.09.003
16. Xu, H.; Dzhashiashvili, Y.; Shah, A.; Kunjamma, R. B.; Weng, Y. L.; Elbaz, B.; Fei, Q.; Jones, J. S.; Li, Y. I.; Zhuang, X.; Ming, G. L.; He, C.; Popko, B. m⁶A mRNA methylation is essential for oligodendrocyte maturation and CNS myelination. Neuron 2020, 105(2), 293–309.e5. https://doi.org/10.1016/j.neuron.2019.12.013.
17. Shi, Z.; Wen, K.; Zou, Z.; Fu, W.; Guo, K.; Samnudin, N. H.; Ruan, X.; Sullere, S.; Wang, S.; Zhang, X.; Thinakaran, G.; He, C.; Zhuang, X. YTHDF1 mediates translational control by m⁶A mRNA methylation in adaptation to environmental challenges. bioRxiv 2024, Preprint. https://doi.org/10.1101/2024.08.07.607063. PMID: 39149343.
18. Widagdo, J., Zhao, Q.Y., Kempen, M.J., Tan, M.C., Ratnu, V.S., Wei, W., … & Bredy, T.W. (2016). Experience-dependent accumulation of N6-methyladenosine in the prefrontal cortex is associated with memory processes in mice. The Journal of Neuroscience, 36(25), 6771–6777. https://doi.org/10.1523/JNEUROSCI.4053-15.2016
19. Li, Z., Weng, H., Su, R., Weng, X., Zuo, Z., Li, C., … & Chen, J. (2017). FTO plays an oncogenic role in acute myeloid leukemia as a N6-methyladenosine RNA demethylase. Cancer Cell, 31(1), 127–141. https://doi.org/10.1016/j.ccell.2016.11.017
20. Barbieri, I., Tzelepis, K., Pandolfini, L., Shi, J., Millán-Zambrano, G., Robson, S.C., … & Kouzarides, T. (2017). Promoter-bound METTL3 maintains myeloid leukaemia by m6A-dependent translation control. Nature, 552(7683), 126–131. https://doi.org/10.1038/nature24678
21. Engel, M., Eggert, C., Kaplick, P.M., Eder, M., Röh, S., Tietze, L., … & Chen, A. (2018). The role of m6A/m-RNA methylation in stress response regulation. Neuron, 99(2), 389–403.e9. https://doi.org/10.1016/j.neuron.2018.07.009
22. Gokhale, N.S., McIntyre, A.B.R., McFadden, M.J., Roder, A.E., Kennedy, E.M., Gandara, J.A., … & Horner, S.M. (2016). N6-Methyladenosine in Flaviviridae viral RNA genomes regulates infection. Cell Host & Microbe, 20(5), 654–665. https://doi.org/10.1016/j.chom.2016.09.015
23. Li, N.; Hui, H.; Bray, B.; Gonzalez, G. M.; Zeller, M.; Anderson, K. G.; Knight, R.; Smith, D.; Wang, Y.; Carlin, A. F.; Rana, T. M. METTL3 regulates viral m⁶A RNA modification and host cell innate immune responses during SARS‑CoV‑2 infection. Cell Rep. 2021, 35(6), 109091. https://doi.org/10.1016/j.celrep.2021.109091. PMID: 33961823
24. Lichinchi, G., Gao, S., Saletore, Y., Gonzalez, G.M., Bansal, V., Wang, Y., … & Rana, T.M. (2016). Dynamics of the human and viral m6A RNA methylomes during HIV-1 infection of T cells. Nature Microbiology, 1, 16011. https://doi.org/10.1038/nmicrobiol.2016.11